I've worked for many years with the good folks at the Embryo Project Encyclopedia (embryo.asu.edu and @embryoproject). They've asked me to do a developmental series for the public.The first series I do will be on a molecular technique called In Situ Hybridization (ISH).
ISH is a technique that biologists use in the lab to visualize the products of gene expression. ISH is very useful for visualizing how genes are regulated during development. But what exactly are we visualizing with ISH?
DNA is a biological molecule that is safely stored and copied in the nucleus. Transcription, or the writing of RNA from a DNA template also occurs in the nucleus. After some modifications are made (like receiving it's 5' cap and poly-AAA tail), the mature mRNA can be transported out of the nucleus into the cytoplasm. The cytoplasm of the cell is where proteins will be made.
For the ISH visualization technique, the goal is to see certain mRNA transcripts that are floating around in the cytoplasm of the cell.
Development is fascinating because common genes are used in different ways, in different places and at different times. Even slight changes in this "temporspatial pattern" can cause drastic changes. Embryos are continually transforming their body via tissue generation, or the formation of body structures, which is driven by gene expression.
ISH is an amazing tool we can use to qualitatively discover where and when genes are expressed.
Take home message:
ISH is a technique that lets us visualize the product of genes, mRNA, in the cell. An antisense probe is used to bind to the mRNA. The probe is designed to target specific gene products, which have been transcribed then transported from the nucleus to the cytoplasm. It is there that the probe will detect mRNA. Anywhere that purple dye is visible is where the probe has bound to this select mRNA.
This
blog will be a 2-parter. The first one will provide images of developmental
gene expression. The next one will focus on the steps in the protocol to
explain why certain techniques are used.
The
following pictures were taken by me, and are the results of experiments I
ran. Not all the embryos were dissected by me (ie mouse embryos), as
previous students may have banked them. Below are the embryos that have been taken through the in situ hybridization protocol.
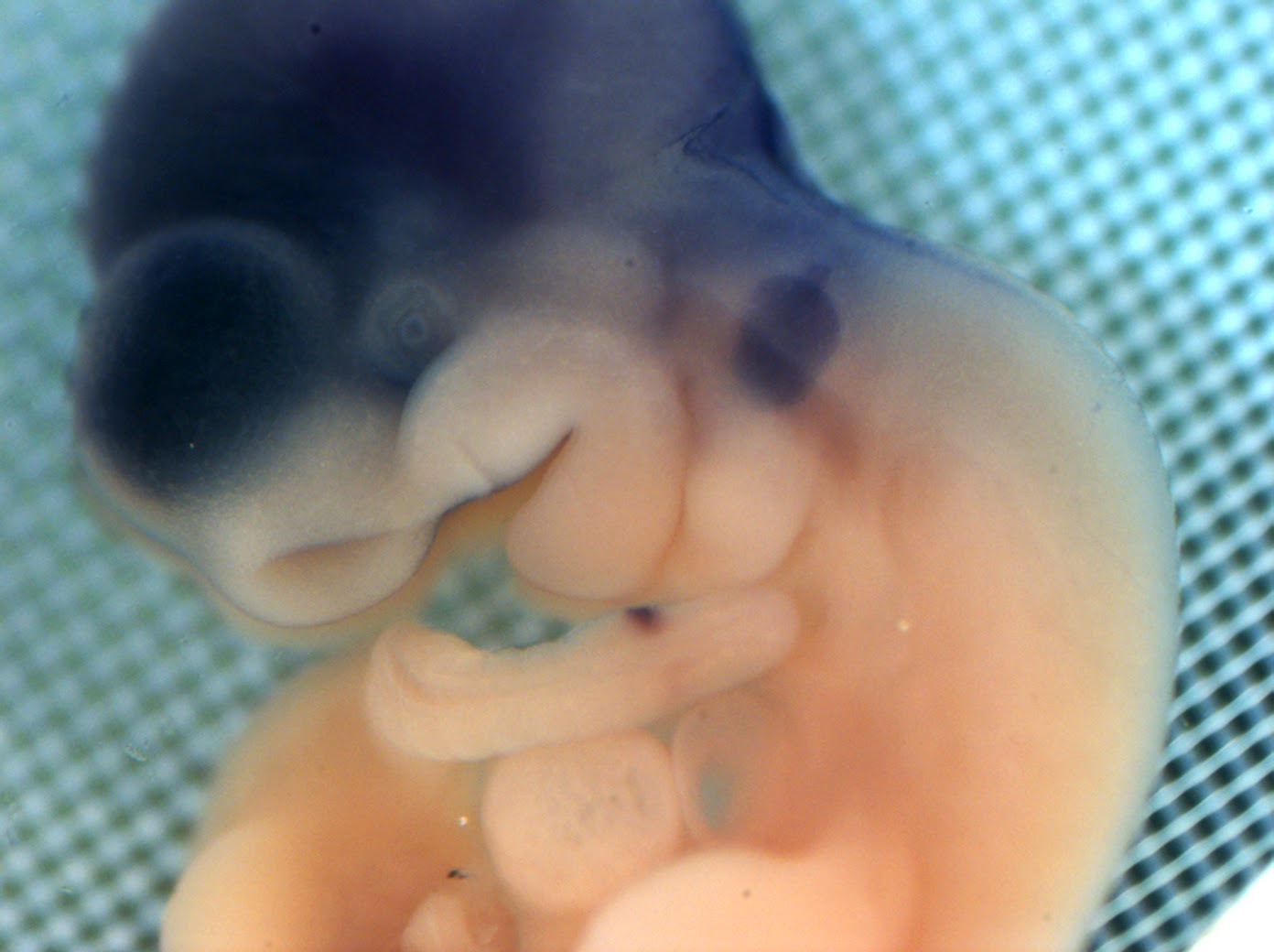
ID2 gene expression
8.5 - 10.5 dpc mouse embryos
 |
| ID2 gene in mouse embryo. Cranial cavity and forming eyes. Top view. |
 |
| ID2 gene expression along future spine. View from below - the dorsal edge of the neural tube is in purple. |
AXIN2 gene expression
8.5 - 10.5 dpc mouse embryos
 |
| Axin2 expression in somitic compartments along the future spine |
 |
| Axin2 gene expression in a mouse embryo. |
GLCC1 gene expression
8.5 - 10.5 dpc mouse embryos
 |
| GLCC1 gene expression in a mouse embryo |
 |
| GLCC1 gene expression in mouse |
ID1 gene expression
8.5 - 10.5 dpc mouse embryos
 |
| ID1 gene expression in a mouse embryo. Side view of a mouse embryo. You can see expression along the somitic edges |
 |
| ID1 gene expression in a mouse embryo |
 |
| ID1 gene expression in a mouse embryo |
 |
| ID1 gene expression in a mouse embryo. Future spine to the left. Side view. |
 |
| ID1 gene expressionn in a mouse embryo Side view. |
 |
| ID1 gene expression in a mouse embryo. View from the back of the head (future hindbrain progenitors in purple) |
 |
| ID1 gene expression in a mouse embryo. View from the future neck, along the spine. Dorsal edges of the neural tube are lined in purple. |
 |
| ID1 gene expresison in a mouse embryo. Side view. |
Lunatic Fringe (lfng) Gene in 9.5 dpc mouse embryos
 |
| Lunatic Fringe gene expression in a mouse embyro. Side view. |
 |
| Lunatic Fringe gene expression in a mouse embyro. Dorsal view. |
 |
| Lunatic fringe gene expression in a mouse embyro |
 |
| Lunatic fringe probe in a mouse embryo |
 |
| Lunatic fringe expression in a mouse embryo. Close up of the tail bud and pre-somitic mesoderm. |
The following pics were taken on an
inverted microscope. The embryos were thin enough to appear translucent.
Below, you can see tiny blocks of mesoderm called somites. They will
form pre-vertebrae and help pattern muscle, bone and cartilage
development. The dark spot is where high levels of lunatic fringe are
being expressed.
In most vertebrates, somites begin to
organize at the anterior pre-somitic mesoderm, progressively forming
into balls as they are displaced anteriorly away from the determination
front. Epithelialization occurs, with cellular changes to the surface
area of the somite creating distinct boundaries. The inner core of the
somite contains more disorganized mesenchymal cells. Cell to cell
interactions are critical for mesoderm formation. Changes to the tissue
type are a product of cell-to-cell interactions as well as the
expression of Eph/Ephrin signaling (Barrios et al., 2003).
 |
| AP axis of a mouse embryo. Taken on the inverted microscope |
Mechanisms
for segmenting tissue is among some of the oldest evolutionary
innovations. Homeobox genes involved in segmentation have also been
found in marine worms, predating invertebrates and vertebrates (Hall
2008). The segmentation mechanism may even predate Animalia, as there is
evidence of plant oscillatory gene expression involved in segmentation
(Richmond and Oates, 2012). Although there is a high variation of the
segmentation clock, there appear to be three conserved features among
plants and animals: cellular oscillations, local synchronization of
cells, and global control of cell cycle arrest (Richmond and Oates,
2012). Many molecular and cellular processes go into forming the segmented blocks of mesoderm, seen in the picture above.
| Taken on the inverted microscope |
| Optic placode/future eye. Taken on the inverted microscope |
The
vertebrate paraxial mesoderm contains very similar gene regulatory
networks that were involved in primitive head segmentation. One of the most important
components for forming the paraxial mesoderm is Wnt signaling in the
posterior tailbud. Wnt keeps the progenitor cells in a state of
proliferation, where they then migrate anteriorly into the pre-somitic
mesoderm. The generation of this raw material for producing somites is
critical for somitogenesis.
| Tailbud. Taken on the inverted microscope |
The
mesoderm is one of four germ layers to give rise to all the tissues and
structures of the body. Mesoderm is created through the process of
gastrulation as cells ingress through the primitive streak and undergo
major multiscale changes, from individual cell morphology to changes in
how cell populations are organized. The movement and rearrangement of
entire cell populations is a common mechanism during development, and
how the movements are orchestrated have changed during chordate
evolution. Intercalation and the formation of compartments are two
examples of such mechanisms.
As
the name suggests, the paraxial mesoderm (PM) runs parallel to the AP
axis along the neural tube and notochord. The PM can be thought of as a
distinct biological object with it's own constituent parts, evolutionary
history and emergent properties. By unraveling the various components
of a modern vertebrate's PM, one is reminded that evolution is
non-directional and is more like a random walk through possible range of
functional outcomes.
Sensitivity
Lunatic fringe is a pretty robust probe.
It's quite easy to get a signal. Thus, to test the sensitivity of my
technique, my PI had me use very small amounts of probe to see if I can
still get a signal. Indeed, even with 0.1 microliters and 50 nanoliters
of probe, I still got a signal. With the 50 nanoliters, I only got
signal on one side of the neural tube.
| Sensitivity testing - only 50 nanoliters of probe used |
| Sensitivity testing - only 0.1 microliter of probe used |
All embryos were treated ethically, according to IUCAC protocol.
Attribution-ShareAlike
CC BY-SA
This license lets others remix, tweak, and build upon your
work even for commercial purposes, as long as they credit you and
license their new creations under the identical terms. This license is
often compared to “copyleft” free and open source software licenses. All
new works based on yours will carry the same license, so any
derivatives will also allow commercial use.
























No comments:
Post a Comment